The intraspecific variation in wing lengths of 10,551 species (birds, moths, butterflies, and cicadas) collapse onto the same universal function under rescaling, allowing for highly accurate trait imputation.
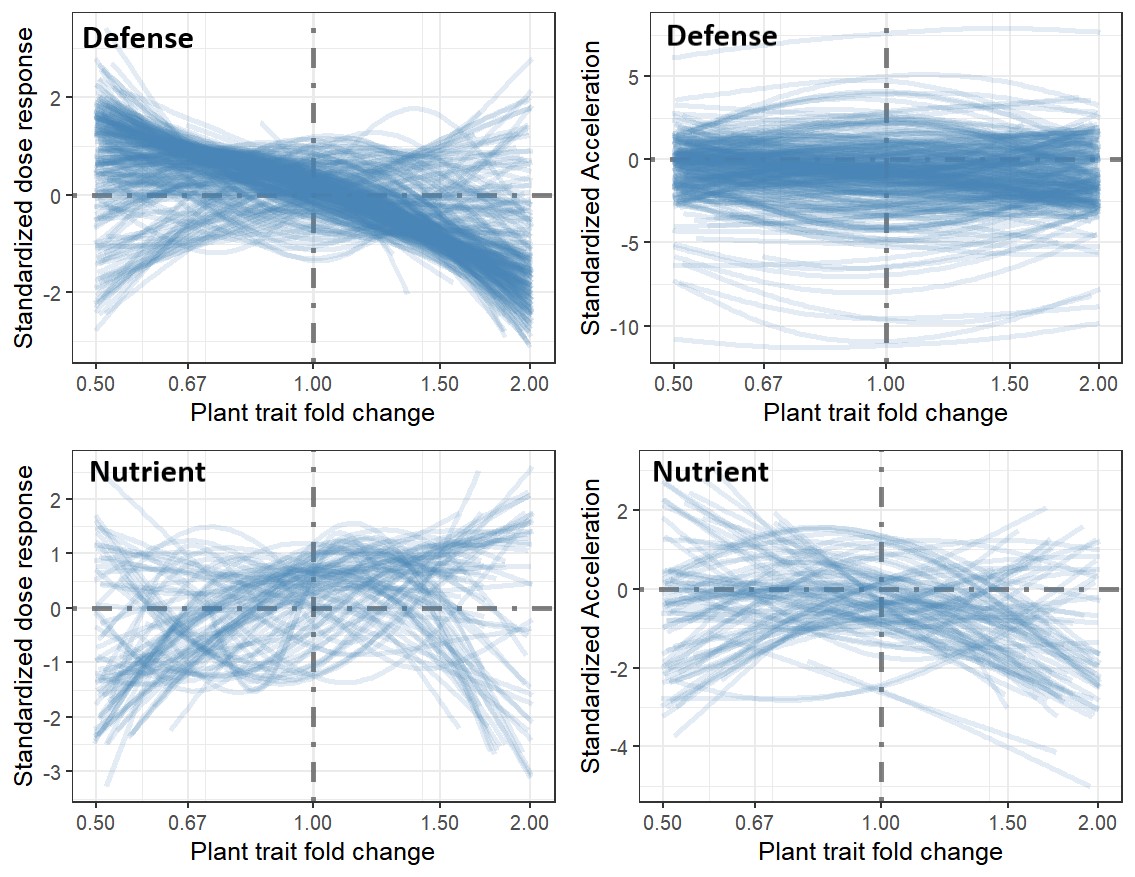
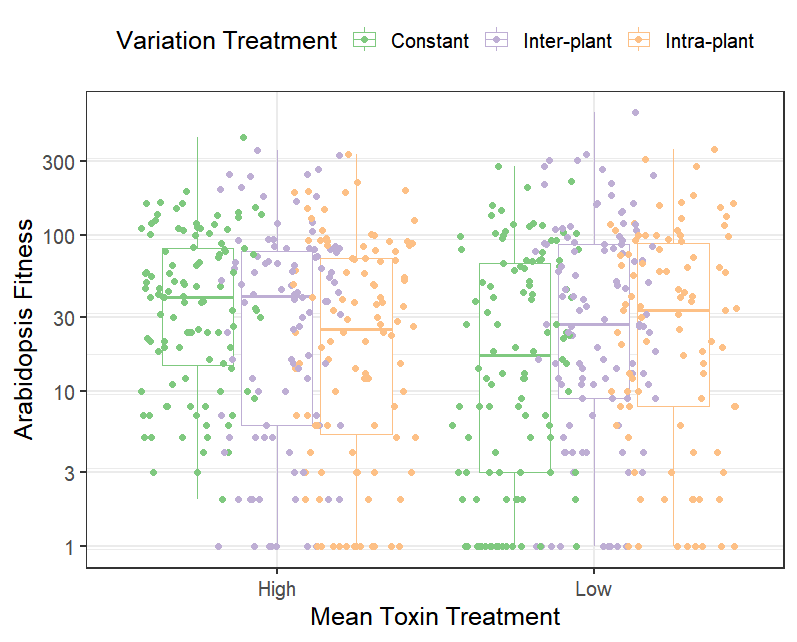
Plant tolerance to herbivory tends to be highly nonlinear which is important for plant population dynamics, community stability, and the existence of alternative stable states in the evolution of mean herbivore damage.
Among plant herbivory within a population in the HerbVar data set show a characteristic strong right skew. Many plants get very little or no herbivory. Deviations from neutral predictions are explained by plant phylogeny.
A running theme of my research is exploring biological questions from a top-down statistical perspective. This approach is motivated by the recognition that nature's complexity often exceeds one's ability to fully understand and measure it at scale. Nevertheless, simple, useful rules may emerge from complex systems. To uncover these general principles or to extrapolate insights to ecologically relevant spatiotemporal scales, one can exploit the separation of scale and statistical properties of the system to generate nontrivial and testable predictions. I work on a variety of topics that generally revolve around plants and insects using this approach. My work tends to be grounded in an attempt to describe patterns in real world data and experimental results.
For instance, I developed a model of herbivory from first principles based on neutral stochastic sampling of herbivores from a regional pool. Comparison of the model to an extensive global data set of herbivory from the HerbVar Network revealed that neutral stochastic processes dominate natural variation in herbivory (Pan & Wetzel 2024). This is important because it suggests that high natural variation in herbivory is an inherent property of the system, mostly comes down to luck, and does not imply that it is driven by variation in plant quality. It is also important for plant fitness because plant tolerance to herbivory tends to be highly nonlinear, so variable herbivory reduces average plant fitness more than expected.
More recent work involves using methods borrowed from statistical physics and economics to model temporal fluctuations and distribution of traits. I am also exploring the potential to model the diversity and occupancy patterns in phytochemistry by modeling the observation process and underlying stochastic gene expression.